Dúvidas sobre
doenças genéticas?
Pergunte que eu respondo!

 A síndrome de Klinefelter é uma condição cromossômica em meninos e homens que pode afetar o desenvolvimento físico e intelectual. Mais comumente, os indivíduos afetados são mais altos do que a média são incapazes de ter filhos biológicos (inférteis); porém os sinais e os sintomas da síndrome de Klinefelter variam entre meninos e homens com essa condição. Em alguns casos, as características da condição são tão leves que a condição não é diagnosticada até a puberdade ou idade adulta, e os pesquisadores acreditam que até 75 por cento dos afetados homens e meninos nunca são diagnosticados.
Como acontece essa síndrome? Dentro das nossas células (no núcleo), o nosso material genético está organizado na forma de cromossomos, onde temos 22 pares de cromossomos autossômicos e 2 denominados sexuais. A síndrome de klinefelter acontece quando há dois cromossomos X no indivíduo fenotipicamente masculino. O cariótipo mais comumente associado a síndrome de Klinefelter é 47 XXY Porém, pode haver alguns indivíduos mosaicos em que algumas células possuem um número adequado de cromossomas mas enquanto outros possuem um X supranumerário e em 90% dos casos o cariótipo da síndrome de Klinefelter é o 47 XXY. Geralmente a Síndrome de Klinefelter ocorre aleatoriamente devido a um erro na não-disjunção meiótica, esse erro pode acontecer tanto na gametogênese materna quanto na paterna. Os sintomas da síndrome da síndrome de Klinefelter geralmente do fenótipo clínico típico é de um homem de alta estatura, membros compridos, pequenos testículos, ginecomastia (aumento das mamas) e diminuição do número de espermatozoides. Também se associa a gravidade do fenótipo com o número de cromossomo X a mais. Em alguns casos a síndrome de Klinefelter só vai ser diagnosticada quando o casal foi procurar um médico para pesquisar infertilidade, sendo a síndrome diagnosticada em 3% dos homens inférteis. Alguns estudos têm demonstrado que o cromossomo X extra e o material que ele codifica é responsável pela hialinização e fibrose dos testículos levando a sintomas de falência que podem aparecer como puberdade atrasada e também como ginecomastia. O diagnóstico da síndrome de klinefelter é confirmado a partir da suspeita clínica pelo cariótipo, ao ser encontrado o cromossomo X a mais em uma linhagem celular ou em todas as células. A síndrome de Klinefelter infelizmente não tem cura, o tratamento é multidisciplinar para as várias complicações que essa síndrome pode ter. Suplementação com testosterona, com supervisão de um endocrinologista pediátrico pode ser usada em alguns casos, principalmente para diminuir as características físicas dessa síndrome, como por exemplo para aumentar o tamanho do pênis e virilização. A longo prazo indivíduos com síndrome de Klinefelter podem vir a desenvolver diabetes tipo 2, dislipidemia e esteatose hepática, além de doenças tromboembólicas, demonstrando a importância do seguimento correto e do diagnóstico. Por: Nicolas Guzman, Acadêmico do curso de Medicina da UNILA. FONTE: https://pubmed.ncbi.nlm.nih.gov/?term=%28Klinefelter+Syndrome%5BMA JR%5D%29+AND+%28Klinefelter+syndrome%5BTI%5D%29+AND+english%5Bla% 5D+AND+human%5Bmh%5D+AND+%22last+1800+days%22%5Bdp%5D
0 Comments
 O desenvolvimento do câncer de mama está relacionado a inúmeros fatores, que deixam o indivíduos mais suscetível a ficar doente, são eles: aumento da idade, fatores endócrinos/história reprodutiva, fatores comportamentais/ambientais e fatores genéticos/hereditários (ADAMI et al., apud INCA, 2021).
No caso do câncer de mama, o histórico familiar constitui uma condição epidemiológica de alto risco, pois indica a existência de uma predisposição hereditária ao câncer de mama que englobam as decorrentes circunstâncias: familiares acometidos em três gerações contínuas; dois ou mais parentes de primeiro grau observados com a doença no ciclo. A ginecomastia, edema do tecido mamário masculino, proveniente de um desequilíbrio hormonal, é um fator de risco para o câncer de mama masculino (VIEIRA et al.; ZEITUNE et al. apud COELHO et al., 2018). Para que uma célula deixe de ser saudável e se transforme em uma célula neoplásica ocorrem diversas mutações alteram a grade da matriz, ou seja, quando há uma desregulação da função de genes que agem indiretamente ou diretamente na proliferação ou na sobrevida das células, como os genes supressores de tumor e proto-oncogenes. Os oncogenes mais comuns são alelos mutantes de uma classe de genes celulares normais. São conhecidos como proto-oncogenes e seu funcionamento ou expressão, quando alterados, resultam em estimulação anormal da divisão celular e proliferação (AMENDOLA; VIEIRA apud COELHO et al., 2018). A presença de apenas um alelo com mutação já pode modificar o fenótipo de uma célula normal para maligno. Em contrapartida, os genes supressores tumorais, como por exemplo os genes BRCA1 e BRCA2, quando estão alterados, ocorre um mecanismo de perda de função de ambos os alelos, ou seja, um alelo mutante é herdado e o segundo alelo é inativado por um evento somático. Ademais, genes supressores tumorais, quando mutados, suprimem os chamados genes protetores (gatekeepers) que são responsáveis por regular diretamente o crescimento celular (DUFLOTH et al.; KERR; ASHWORTH; MACLEOD; NUSSBAUM; MCINNES; WILLARD apud COELHO et al., 2018). A predisposição genética a tumores é mediada pela herança da inativação de genes supressores de tumores, particularmente em famílias de alto risco. Os mais importantes genes supressores de tumor associados com o câncer de mama são os genes BRCA1 e BRCA2. Mulheres portadoras de mutações no BRCA1 possuem até 80% de chance de desenvolver câncer de mama, e até 60% de desenvolver câncer de ovário em sua vida (INCA; DUFLOTH apud COELHO et al., 2018) O gene BRCA1 está localizado no braço longo do cromossomo 17, constituído por 22 éxons codificantes, e codifica para uma proteína de 1.863 aminoácidos. Enquanto o gene BRCA2 se encontra no braço longo do cromossomo 13, sendo composto de 27 éxons codificantes, dos quais 26 codificam uma proteína com 3.418 aminoácidos. Esses dois genes são incumbidos de codificar proteínas nucleares expressas que são conservadas com integridade genômica por regular o reparo de DNA. Sua função é coibir o desenvolvimento de tumores por meio do reparo de DNA que estão alterados. Assim, as proteínas que são codificadas pelos genes interagem com outras proteínas para reparar as quebras de DNA. As proteínas BRCA1 e BRCA2 também são responsáveis por organizar a atividade de outros genes e também exercem um papel importante para o desenvolvimento embrionário (ECONOMOPOULOU; DIMITRIADS; PSYRRI; CARDOSO; FAGANELLO; LAJUS; AMENDOLA; VIEIRA apud COELHO et al., 2018). Os fatores genéticos/hereditários foram relacionados também à presença de mutações em outros genes como: PALB2, CHEK2, BARD1, ATM, RAD51C, RAD51D e TP53. Mulheres que possuem vários casos de câncer de mama e/ou pelo menos um caso de câncer de ovário em parentes consanguíneos, sobretudo em idade jovem, ou câncer de mama em homem também em parente consanguíneo, podem ter predisposição hereditária e são consideradas de risco elevado para a doença. O câncer de mama de caráter hereditário corresponde, por sua vez, a apenas 5% a 10% do total de casos (BREAST CANCER ASSOCIATION CONSORTIUM; GARBER et al.; ADAMI et al., apud INCA, 2021). Por: Jéssica Albino, Enfermeira e Acadêmica do Curso de Medicina da UNILA. REFERÊNCIAS BRASIL. Instituto Nacional do Câncer - Inca. Ministério da Saúde. Controle do câncer de mama: fatores de risco. 2021. Disponível em: https://www.inca.gov.br/controle-do-cancer-de-mama/fatores-de-risco. Acesso em: 14 out. 2021. COELHO, Aline Silva; SANTOS, Marielle Anália da Silva; CAETANO, Rosecleide Inácio; PIOVESAN, Camila Fátima; FIUZA, Larissa Aparecida; MACHADO, Ricardo Luiz Dantas; FURINI, Adriana Antônia da Cruz. Hereditary predisposition to breast cancer and its relation to the BRCA1 and BRCA2 genes: literature review. Revista Brasileira de Análises Clínicas, [S.L.], v. 50, n. 1, p. 1-7, abr. 2018. Revista Brasileira de Analises Clinicas. http://dx.doi.org/10.21877/2448-3877.201800615. 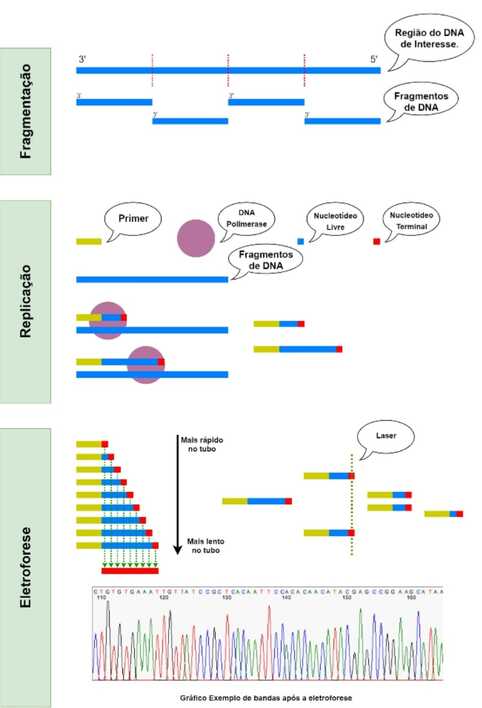 Essa postagem tem por intuito ser a primeira de uma série de 3 postagens falando sobre técnicas de sequenciamento genético. Neste primeiro artigo é abordado os eventos que ocorrem durante o sequenciamento usando o Método de Sanger.
Com os avanços dos conhecimentos sobre o material genético provocados pelas descobertas de Watson e Crick, ao final do século XX surgiu a necessidade de compreender conceitos além do “formato” da molécula carregadora de informações. Foi então em 1977 que Frederick Sanger e seus associados desenvolveram um método capaz de encontrar a ordem de pares de bases para um certo fragmento de uma molécula de DNA. Essa nova tecnologia de sequenciamento genético (o “Sequenciamento Sanger”) era única no mercado e, consequentemente, se tornou a com o mais amplo uso na atualidade. Para aplicação do Método de Sanger é preciso, primeiro, realizar uma preparação da amostra, que é a região do DNA de interesse a ser sequenciado. Esse preparo consiste em três partes principais: a fragmentação, a multiplicação e o isolamento. Na parte de fragmentação, a amostra é fragmentada em partes menores, as quais serão protagonistas durante o sequenciamento. Esses pequenos fragmentos, então, são separados e multiplicados, no intuito de aumentar a quantidade de material amostral. Um dos processos de multiplicação mais usados é a inserção destes fragmentos em plasmídeos de bactérias, as quais vão se proliferar através de clonagem. Por fim, após apresentar-se a quantidade de material desejada, é necessário isolar o material genético dos outros componentes usados para a fragmentação e proliferação. Dessa forma, cada secção da molécula de DNA apresenta, agora, em grande quantidade; obtendo-se, assim, amostras prontas para a análise. Após o preparo, uma das amostras (um dos grupos de fragmentos da molécula de DNA maior) é inserida em um ambiente rico em nucleotídeos e componentes para replicação (primers e DNA Polimerases). Por haver vários componentes para a replicação e pequenos fragmentos idênticos de DNA, inicia-se o processo de replicação. No entanto, ocorre a presença de dois tipos de nucleotídeos: os livres, que são nucleotídeos comuns, semelhantes aos usados em uma replicação em condições normais; e os marcados com material fluorescente, que, além da presença desse material, também apresenta uma alteração na ribose (substituição dos grupos hidroxila por hidrogênios) que permite com que ele não possa realizar ligação fosfodiéster, o que permite que sejam chamados de nucleotídeos terminais. Semelhante ao PCR, ciclos de temperatura são usados para a realização do processo de sequenciamento. Inicia-se com um aumento de temperatura para separar as duas moléculas de DNA da fita dupla durante o preparo da amostra. Já em uma temperatura menor, ocorre a aproximação do primer e da DNA Polimerase. Nesse momento, são depositados nucleotídeos na sequência adequada para o fragmento sendo analisado, no entanto de forma aleatória entre os nucleotídeos livres e terminais. Ao ser depositado um nucleotídeo terminal, a DNA Polimerase para de realizar a replicação, já que não é possível continuar o processo sem continuar fazendo ligações fosfodiéster. Dessa forma, a partir do fragmento analisado, fragmentos menores e de diferentes tamanhos são formados, com seus nucleotídeos finais estando marcados pelo material fluorescente. Então, é realizado uma eletroforese com o conteúdo resultante das reações da replicação. A eletroforese consiste na passagem dessas moléculas por um tubo com gel a partir de movimentos eletromagnéticos, devido a diferença de potencial entre as extremidades do tubo. Também, em um ponto mais distante no percurso do tubo, é introduzido um Laser capaz de estimular a marca fluorescente dos nucleotídeos terminais. Nesse ponto, devido ao atrito entre as moléculas de DNA e o gel, as moléculas menores percorrem a extensão do tubo mais rapidamente, ficando a frente, enquanto as mais longas ficam por último – ativando, assim, o laser em tempos diferentes. Portanto, uma câmera capta a luz resultante da marca dos nucleotídeos terminais, a qual se diferencia entre ATCG através de cores – como A sendo verde, T azul, C vermelho e G amarelo, por exemplo. É por conta da diferença de tamanho que é possível determinar a ordem dos nucleotídeos terminais que durante a eletroforese. Isso porque, uma molécula de DNA com um primer e um nucleotídeo terminal, ocuparia a vanguarda do percurso do tubo; sendo, assim, o nucleotídeo correspondente ao primeiro da sequência do DNA analisado. O segundo, seria um nucleotídeo mais longo (apresentando um primer, um nucleotídeo polimerizado do tipo livre e outro nucleotídeo polimerizado do tipo terminal), o terceiro 2 mais longo e assim sucessivamente. Isso permite, portanto, determinar a ordem de nucleotídeos em uma molécula de DNA. Por: Freddy Romanno Aires Nader, Acadêmico do curso de Medicina da UNILA |
